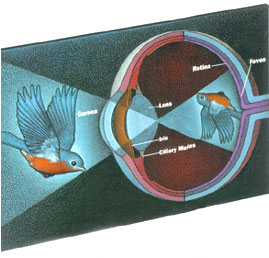
Neurologie: neurale netwerken, patroonherkenningDe term "neurale netwerken" is tegenwoordig voornamelijk bekend uit de techniek, als software die in staat is van ingevoerde gegevens te leren - om daar patronen in te herkennen. De term wijst natuurlijk op de biologische oorsprong van het proces, in wat er gebeurt in de hersenen, of netter: het zenuwstelsel.In dat zenuwstelsel zijn vele vormen van neurale netwerken, met als mogelijke eerste tweedeling die tussen min-of-meer bolvormige en min-of-meer laagvormige. Het proces genaamd "patroonherkenning" vindt plaats in laagvormige netwerken, het meest duidelijk en vermoedelijk evolutionair ook het eerst in het oog, preciezer: het netvlies, zie de illustratie (van harunyahya.com  ): ):
Netvlies aan de achterkant van de oogbol links en in detail rechts. Hier wordt een mogelijk evolutionair pad van de ontwikkeling van neurale netwerken geschetst aan de hand van dit voorbeeld. De vermoedelijke eerste soort neuronen verbonden waarnemingsorganen direct met ledematen dat wil zeggen spieren (illustratie van hier ):
Met het waarnemingsorgaan verbonden aan de rechterkant en spier aan de linker. Dat heeft zich ontwikkeld in vele soorten neuronen met gespecialiseerde functies. Die aan de waarnemingsorganen en naar de spieren zitten voornamelijk in de door het gehele lichaam lopende zenuwbundels (12 stuks). Maar het overgrote deel is zogenaamde interneuronen, die andere neuronen verbinden. Hier gaan we dus kijken naar het oog. Het nu bekende oog heeft miljoenen lichtgevoelige cellen, maar in principe is dat begonnen met eentje. Wat niet zo moeilijk was, want veel van de werkzame stoffen in een cel zijn lichtgevoelig. Dat wil zeggen: onder invloed van licht vallen ze uit elkaar, zoals vele kleurstoffen dat doen: ze vervagen onder het (zon)licht. Het uit elkaar vallen van een bepaalde stof wordt in de cel gedetecteerd, en daar is een lichtgevoelige cel. Het tweede element is een neuron dat het signaal van de lichtgevoelige cel doorgeeft aan de rest van het lichaam. Waar en wat dan ook moge zijn. Dus bijvoorbeeld spieren. Het is duidelijk dat meerdere lichtgevoelige cellen meer informatie oplevert dus overlevingsvoordeel, zodat er ogen ontstaan - zie een voorbeeld van een evolutiemodel (illustratie van hier ):
Maar de basale stap is natuurlijk van één naar twee. Overgaand op schematische tekeningen, krijg je dan iets als onder (lichtcel rechts, neuron links):
De stap naar twee lichtcellen faciliteert de eerste vorm van patroonherkenning. Want stel dat het gaat om "meer of minder licht" - dan zijn er logisch gezien vier mogelijkheden: 1 detecteert licht en 2 detecteert licht, 1 wel en 2 niet, 1 niet en 2 wel, of allebei niet. De koppeling met evolutie is deze: stel de lichtgevoelige cellen zitten naast elkaar op een plek van het lichaam van het wezen. Als dat wezen zelf beweegt (of het onderdeel waar de cellen zitten), zal vermoedelijk beide cellen meer, of beide cellen minder licht ontvangen. Als het wezen stilligt en er beweegt iets buiten het wezen met gevolgen voor de hoeveelheid licht, is er een goede kans dat de ene lichtgevoelige cel daar eerder iets van meekrijgt dan de andere. De menselijk-verbale interpretatie luidt: "beweging". En dus mogelijk een ander wezen. En dus mogelijk gevaar. En dus is het het beste om het zekere voor het onzekere te nemen, en weg te wezen. En daar is het evolutionaire verschil: alletwee de lichtgevoelige cellen licht(er), of alletwee donker(der), heeft een andere betekenis dan één licht(er) en één donker(der). Maar daarvoor moet er wel "ergens" bekend zijn wat beide cellen waarnemen - oftewel: de betreffende neuronen moeten verbonden zijn met beide cellen. Omdat de natuur moet blijven functioneren "tijdens de verbouwing", worden nieuwe zaken altijd gebouwd op oude. Dat wil hier zeggen: voor de neuronen die signaaltechnisch verbonden zijn met beide lichtcellen wordt een nieuw stel neuronen gebruikt. Gevende dit schema (signalen van rechts naar links):
Waarom twee stuks verbonden neuronen? Dat doet de natuur (bijna) altijd: twee kanten van de zaak nemen. Dus een neuron dat de al-dan-niet symmetrische (aan-aan, uit-uit) en een neuron dat de asymmetrische kant (aan-uit, uit-aan) voor zijn rekening neemt. Rest de vraag: hoe zit de logica in elkaar om dit resultaat te krijgen? Dat is uiterst simpel: zet tussen één van de kruisverbanden een omkering (correcter: één van de andere drie is ook goed). In dit soort schema's aan te geven met een minteken erbij. Zodat het schema wordt:
Ga zelf de uitkomsten na, maar neem niet het gebruikelijke 1 en 0, maar 1 en -1 voor respectievelijk "licht" en "donker" (weer zo'n natuur-ding: die werkt nooit met "niets", ook niet als "niets" moet worden aangegeven). Twee bezwaren: hoe moet een neuron een signalen omkeren? Dat is weer dezelfde "natuur"-kwestie: neuronen hebben niet één maar twee soorten primair signaal (twee primaire neurotransmitters Tweede bezwaar: de uitkomsten zijn dubbel: +2 en -2 . Klopt. Maar de neuronen werken niet met enkelvoudige, statische, signalen, maar met pulspatronen - het herhaaldelijk en voortdurend afvuren waarbij het gaat om de frequentie. Het minteken betekent dan alleen dat het pulspatroon een stukje verschoven is: piek wordt dal en dal wordt piek. Het teken (de "fase") is onbelangrijk. Kenners van de digitale signaalverwerking zullen het schema van de laatste figuur onmiddellijk herkennen: dit is de basis van het procedé genaamd "Fast Fourier Transformation" of FFT (Wikipedia) - welk basis-schema om inzichtelijke
redenen de "butterfly" of "vlinder" wordt genoemd.
Een volledig FFT-schema gebruikt meerdere van deze basis-vlinders naast
elkaar, in verband het snelheid van berekening het liefst in een exponent van
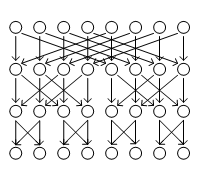
2: 2, 4, 8, 16 enzovoort. Hier het voorbeeld van acht stuks en drie lagen:
De algemene functie van zo'n soort schakeling heet Fourier-analyse Hier is een anatomische en schematische weergave van het netvlies achter het oog (van Webvision
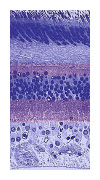
De daadwerkelijke microscopische opname laat al een duidelijke structuur in lagen laat zien, met afwisselend lagen schakelementen, de neuronen, met daartussen lagen met kruislingse verbindingen. Bijna geheel volgens het schema van de FFT-analysator. Het grote verschil is de afname van het aantal neuronen in de lagen van boven naar beneden. Van ingang naar uitgang. Dit is de simpelste vorm van een (niet-triviaal) neuraal netwerk En bij het netvlies is bekend wat het doet: het vormt het puntje-voor-puntje beeld van de losse lichtgevoelige cellen om in meer algemene concepten als lijnen, vlakken, enzovoort. Met twee uiterst belangrijke fysiologische functies: het aantal uitgangen wordt sterk verminderd, die als een (dunne) bundel naar het brein gaan. En als tweede: de contouren gaan sneller dan de vlakken - want bevattende voor het overleven belangrijkere informatie. Omdat het netvlies zo'n basale functie vervult direct achter een primair waarnemingsorgaan is het aannemelijk dat het het eerste neurale netwerk is. Met ook een speciale vorm, omdat de ingang ervan, het punten-raster, uniek is. In vrijwel alle andere toepassingen komt de ingangsinformatie ook in de vorm van een bundel verbindingen - volgens het algemene schema:
Waarbij de min-of-meer bolvormige structuur van het netwerk zelf komt omdat de neuronkernen in de laag "dikker" zijn dan de losse draden: de axonen. Het eerste grotere neurale netwerk in het centrale zenuwstelsel zit in het midden van de hersenstam en heet olive of olivary nucleus, zie onder:
Helder getekend is de donker ingekleurde laag met daarin de neuronale deellagen. Waarvan kennelijk zo veel nodig is dat het in lussen is gaan liggen. Hier niet getekend zijn de in- en uitgangsbundels - in het schematische overzicht op ongeveer gelijke hoogte in de hersenstam staat er een enkele:
De linker vertoont hier een ingangsbundel komende van een kern in de hersenstam, en de rechter een uitgang die via de bundels met nummer 8 naar het cerebellum, de kleine hersenen, gaat. Dit patroon herhaalt zich een aantal malen, zoals te zien hier:
Rechtsonder de (inferior of onderste) olivary nucleus - de directe verbinding met het cerebellum uit het vorige schema is hier weggelaten ten faveure van andere. Het cerebellum Oftewel: de functie van een neuraal netwerk wordt bepaald door de structuur van zijn verbindingslagen Merk op: de in het midden van het cerebellum getekende structuur is de dentate nucleus, die dient als uitgang van het cerebellum. Of deze ook een neuraal netwerk-functie heeft is onbekend. En rechtsboven staat, schematisch en sterk verkleind, het grootste neurale netwerk van het brein: de cortex. Merk ook dit op: in het verbindingenpad tussen olivary nucleus en cerebellum en de cortex liggen nog andere structuren, hier getekend de red nucleus en de thalamus. Deze hebben ook grote hoeveelheden neuronen, natuurlijk, maar duidelijk geen lagenstructuur zoals kenmerkend voor een neural netwerk. Deze organen combineren vermoedelijk wel informatie van meerdere bronnen, maar op een direct schakelende manier. Naar Neurologie, organisatie
|